Introduction
Among biologists, “design” refers to a peculiar coherence between a living structure and a function it performs (Turner, 2007). Bones, for example, are exquisitely constructed cantilevers, built to bear their loads with an elegant economy of form and materials (Currey, 1984), and it seems perfectly apt to say that bones are well-designed. Indeed, to describe them in any other way seems pedantic. Awkwardness attaches to the word, though, because “design” readily conjures up notions of a designer, that troublesome Master Craftsman that Plato introduced in his Timaeus, long the mainstay of natural theology, and that serves that purpose still for the resurgent “natural deism” that imbues Intelligent Design theory.
Darwinism convincingly undercuts this type of thinking. Nevertheless, Darwinism is neither finished product nor holy writ, and there remain interesting and unresolved philosophical issues at its core. The most interesting is this very question of biological design: precisely why are living things so aptly constructed for the things they do? Darwinism, at least in its Neodarwinist conception, puts forth what is essentially an atomist answer to the question: biological design arises solely from the interplay of “atoms of heredity” in gene pools, converging over time onto well-functioning phenotypes through natural selection of particular well-function-phenotype-specifying genes. As in classical atomism, biological design is only apparent, and cannot be informed by any broader purposefulness (Dawkins, 1986).
Standing in contrast to Neodarwinism’s rather bleak vision is an older tradition in biology that puts the organism, not the gene, as the central fact of life. The unresolved issue of design rests in that vast terrain of physiology that separates genes from the well-functioning biological machines that mediate fitness. Much of this territory is terra incognita, but what is known about it exhibits a striking purposefulness, which today we call homeostasis, that seems quite at odds with the supposedly purposeless process that gives rise to it. This poses an interesting question for the atomist-minded Neodarwinist: is homeostasis “mere phenotype”, the product of “homeostasis genes” that confer survival advantages? Or is it the other way round: is homeostasis the prime virtue that allows particular genes to thrive? The question is not trivial: In the one instance, evolution is really what Neodarwinism says it is: the playground of whimsical genes that have no purpose other than their own replication. In the other, evolution has an underlying purposefulness and intentionality that is not inherent in the purposeless genes that supposedly run the show.
Complexity, at least as I understand its scientific definition, is cut from the same atomist cloth as Neodarwinism. Both share a goal of deriving emergent phenomena—adaptation and design in the one case, complex function and structure in the other—from simple rules of interaction among myriad low-level components. However, complexity is also prone to the same philosophical quandary as Neodarwinism: what’s running the show? Is emergent complexity, like biological design, simply a “bottom-up” phenomenon, or is there a higher-level organizing principle at work?
I will argue in this paper that any Neoatomist conception of living systems, whether it be Neodarwinism or Complexity Science, or indeed any form of reductionism, will be inadequate to the task, largely because it cannot grasp, even admit, that there is an essential organizing principle—homeostasis—that cannot be derived reductively. I will say at the outset that I am not a practitioner of Complexity Science. I have, however, spent several years studying and thinking about a living system that is often cited as one of the more compelling examples of a complex emergent system: the social insect “superorganism.”
The Macrotermes superorganism
I study the colonial physiology of termites, specifically those belonging to an advanced termite family, the Macrotermitinae. This grouping comprises fourteen genera, distributed into roughly 350 species. All have in common the cultivation of symbiotic fungi as an adjunct to these termites’ normal intestinal digestion of cellulose (Batra & Batra, 1979; Wood & Thomas, 1989; van der Westhuizen & Eicker, 1991). Two of the genera, Macrotermes
Figure?1
A mound built by Macrotermes michaelseni, in northern Namibia. Two of my students are in the foreground: Wendy Park (l) and Grace Shihepo (r).
{kind=link}
Figure?2
A cross section through a nest of a colony of Macrotermes michaelseni. The light-colored bodies are the fungus combs, where the symbiotic fungus is cultivated.
{kind=link}
Figure?3
Plaster filled casts of the internal network of tunnels in Macrotermes mounds. Left: the vertically oriented network surrounding the nest, which is situated in the center. Middle: the large vertically-oriented tunnels in the center of a mound. Right: a partly exposed cast of tunnels situated just beneath the mound surface. Note the numerous egress tunnels projecting to the surface. Mound casts were done in collaboration with Dr Rupert Soar of Loughborough University.
{kind=link}
The mound-building habit is not unique to the Macrotermitinae, but the use to which these termites put their mounds is. Most termites that build mounds use them as nests, that is, as structures to house the colony: the many sterile workers, the queen and king, and the fertile nymphs (also called alates) that will serve ultimately as the colony’s propagules. The Macrotermes mound is not a nest, however—few termites, save for occasional patrolling workers, are found there. The nest itself is a compact subterranean structure that is situated below the mound (Figure 2), housing both the colony’s complement of termites, and the colony’s culture of symbiotic fungi. The mound’s internal architecture departs significantly from the typical architecture of the termite nest. Where the nest tends to an architecture of horizontal galleries connected with small portals, the mound is permeated with an extensive reticulum of large-calibre tunnels with a striking vertical bias (Figure 3). These ramify through the mound, eventually opening to the surface through a number of tiny egress channels.
The egress channels serve two functions. First, they are the principal sites of mound growth. The mound is built by a net translocation of soil by termites from the mound interior and from deep soil horizons to the mound surface: the egress tunnels provide access to the mound surface for soil-carrying termites. The egress tunnels are also zones of porosity in the mound’s seemingly solid surface, which serve the mound’s principal function as an organ of colonial physiology: capturing wind to power ventilation of the underground nest. Because the mound projects upward through the surface boundary layer, it intercepts wind and converts its kinetic energy into a complex field of pressure over the mound surface (Turner 2000, 2001). Via the porous egress channels, this external pressure field drives a complicated flow of air through the mound’s internal network of tunnels, ultimately ventilating the nest.
The need for nest ventilation is acute. The Macrotermes nest is a focus of high metabolic power, which requires a commensurably high collective demand for oxygen, roughly equivalent to that of a mammal the size of a goat. Without ventilation, the nest’s inhabitants would suffocate (Darlington et al., 1997). Remarkably, most of the nest’s collective oxygen consumption is attributable not to the termites but to the cultivated fungi.
This association of termites, fungal symbionts and sophisticated mound architecture displays a remarkable integrity. The termites provide the cultivated fungi an environment that is rich in nutrients and shielded from their principal fungal competitors (Batra & Batra, 1967; Batra, 1971). The fungi, for their part, serve essentially as an accessory digestive system for the colony, composting the hard-to-digest woody material brought back to the nest into a more easily digestible food for their termite hosts. The mound, meanwhile, serves the respiratory needs of both termites and fungi. By any conceivable definition of the word, this makes the entire assemblage a superorganism. By my understanding of the phrase, it also makes it a complex system. To the question I posed above—are ‘bottom-up’ interactions among the agents of this system sufficient to explain this striking organismal integrity?—I will argue that they can, but only up to a point. The remainder of the explanation, I shall suggest, lies in a concept that is largely lost in the reductionist and atomist mind-set of much of modern biology, complexity science included. That concept is homeostasis.
Homeostasis of structure and function in the Macrotermes Superorganism
The concept of homeostasis has its origins with the great French physiologist Claude Bernard, who noted “la fixité du milieu intérieur”—the defense of internal environments—as a basic principle of living systems. It is also a concept of great subtlety and profound implication, on a par with the Second Law of Thermodynamics, which makes homeostasis—like the Second Law—a widely misunderstood, and therefore often abused idea. Usually, the word is used to describe a generalized tendency to steadiness of particular properties of living systems, like body temperature, blood acidity, and so forth. That usage itself is benign, but abuse intrudes when the word is employed without reference to the mechanisms that give rise to the steadiness. One often finds, for example, mere steadiness of body temperature being described as temperature homeostasis. By this logic, the steady temperature inside a very large rock is as good a demonstration of homeostasis as the steady temperature inside a very large elephant. This absurd conclusion is avoided only through an appreciation of the different paths to steadiness of temperature. The same cautionary tale is applicable to many other “dynamically-stable” systems, like buffered chemical reaction pathways, which also tend toward stability, and even exhibit a sort of resistance to perturbation, but only because the “steady state” represents a thermodynamically stable state, such as a local minimum on a reaction space.
Homeostasis is properly understood as a regulated dynamic disequilibrium, held far out of thermodynamic stability and sustained by the active and purposeful management of fluxes of matter and energy between environments. Body temperature regulation provides a useful illustration of this concept. A warm body in a cold environment represents a disequilibrium in potential energy between environments—body and surroundings—that can drive a physical loss of heat from the body. The rate of loss is proportionate to the magnitude of the disequilibrium: hence, colder environments elicit greater rates of heat loss from the body than do warmer environments. This applies to both the hot elephant and the hot rock. Temperature homeostasis can only occur, however, if these physical heat losses are supplanted by the expenditure of metabolic work, in this instance, directed to the production of heat: thermogenesis. Furthermore, thermogenesis must not only occur, but must proceed at a rate that is actively matched to the rates of physical heat loss. Thus, homeostasis is essentially a phenomenon of fluxes: physical fluxes down thermodynamic gradients in potential energy being offset by metabolic work to drive fluxes of matter and energy against these thermodynamic gradients. Of the two hot bodies, only the elephant, and not the rock, is capable of this trick.
This definition of homeostasis can be readily applied to the Macrotermes superorganism, albeit not for temperature: the nest temperature happens to be steady but because it is buffered, not regulated. In this instance, the relevant disequilibrium is in the composition of the nest air with respect to the outside air: nest air is hypoxic, (nest pO2 is roughly 2 kPa less than atmospheric), slightly hypercarbic (nest pCO2 is elevated roughly 2 kPa above atmospheric) and very humid (Turner, 2000, 2001). These disequilibria are established by the nest’s metabolic work rate, which I shall call the metabolic demand. The demand-induced disequilibria also drive physical fluxes of these respiratory gases across the porous boundary of the mound surface: I shall call this the ventilatory flux. The composition of the nest atmosphere is the consequence of a balance between the nest’s metabolic demand, and the mound’s ventilatory flux. Homeostasis of the nest atmosphere occurs when the two fluxes are matched, which appears to be the norm. We know, for example, that mound size is a reliable indicator of the nest’s metabolic demand. More populous nests (which comprise more engines of soil transport in the form of workers) tend to build larger mounds, and more populous nests have higher overall
Figure?4
Partial pressures of oxygen in the nests of three size classes of Macrotermes michaelseni. Despite the large variation of metabolic demand this size variation represents, the concentrations of oxygen in the nest are the same for the three size classes. After Turner (2000).
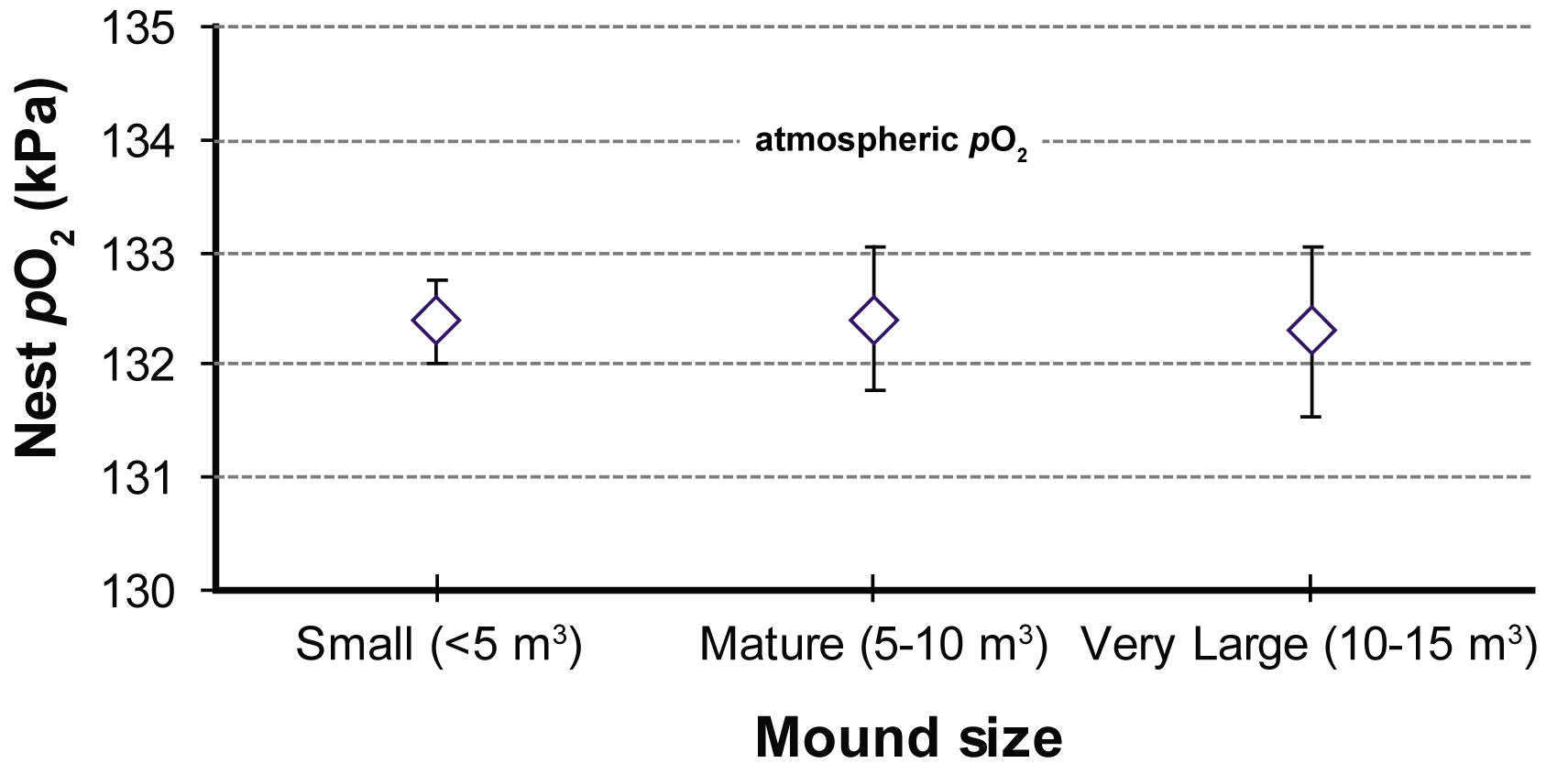{kind=link}
Remarkably, the structure that mediates nest homeostasis—the mound—itself fits the criteria for homeostasis. The mound is a structure in disequilibrium with respect to gravity, maintained by a balance between two fluxes of soil. On the one hand, there is a physical flux of soil from the mound onto the ground surface, driven by erosion by wind or rain coupled to gravity. This flux of soil is substantial, and can amount to several hundred kilograms annually. The mound’s structural disequilibrium is sustained because these physical losses of soil are offset by termites doing work to carry soil up into the mound, out through the egress channels, and depositing it ultimately to the mound surface. The mound’s architecture is therefore a dynamic disequilibrium maintained by two soil fluxes, not so much a structure as a process, an embodiment of two opposing soil movements.
As such, the mound’s architecture can be regulated, just as the nest atmosphere can be. This can be shown dramatically by performing a “complete moundectomy” on a colony, scraping away the mound with a front-end loader (Figure 5). Because this procedure
Figure?5
Rebuilding of a mound following a complete moundectomy. Top: Mound prior to the moundectomy. Middle: The same mound (photographed from a different angle) following the moundectomy. Bottom: The rebuilt mound roughly five weeks later.
{kind=link}
The mound’s architectural regulation is also evident at a less dramatic scale. Mounds often are subject to injury, such as a breach in the porous surface wrought by animals or erosion. This injury elicits a large-scale rebuilding project to repair the breach and restore the mound to its structure prior to the injury. The project proceeds in three stages (Figure 6). The first, or recruitment, stage begins within minutes of the breach, and involves a mobilization of workers from the nest into the mound. The mobilization is elicited by disturbance of the nest atmosphere, wrought by the sudden admission of turbulent wind energy into the mound that accompanies a breach.
The recruitment phase lasts for roughly an hour, and merges into the second, or stigmergic building phase. Stigmergy (literally, “driven by the mark”) is a self-organized building process (Stuart, 1972). A termite lays down a grain of soil onto a surface and cements it into place with salivary glue containing an attractive pheromone: the pheromone-laden grain of soil is the “mark.” Other termites are attracted to this mark, and are driven to deposit new grains of soil onto it. Each new grain, in its turn, is accompanied by another dollop of the attractive pheromone, which produces a still more powerful enticement to other termites to deposit their grains of soil there. On a larger scale, this autocatalytic process produces an organized translocation of soil to form orderly structures, either pillars or sheets initially (Courtois & Heymans, 1991), merging over the longer term into a complex space-filling architecture called a spongy build (Figure 7).
Following an injury, stigmergic building is initiated widely throughout the mound (Figure 6), apparently elicited by turbulent transients within the breached mound. The most intense transients occur near the breach, of course, and these elicit the most intense stigmergic
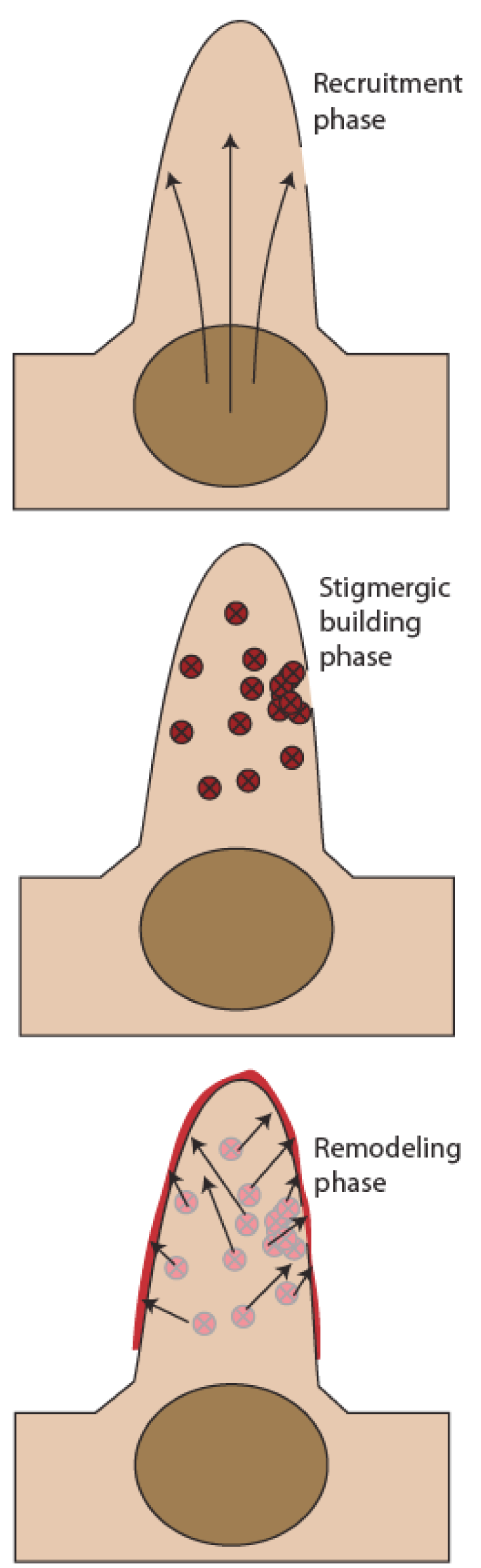{kind=link}
Figure?7
Phases of the mound repair process. Stigmergic building begins with deposition of small piles of soil around the mound’s tunnels (a), which grow initially into pillars and walls (b, c), and eventually into the space-filling spongy build (d, spongy build in a 110 mm PVC pipe; e, plaster cast of the air spaces in a spongy build). Eventually the spongy build is excavated and smoothed to form large smooth-walled spaces (f).
{kind=link}
The system then enters the final remodeling phase, which plays out over the space of several weeks, and involves restructuring the disparate sites of building that were laid down during the stigmergic building phase. This appears to be driven by an aversion of termites to abrupt edges, which are produced abundantly during stigmergic building. When termites encounter an abrupt edge, they dismantle it, vectoring the soil outward through the egress channels and ultimately to the mound surface. The remodeling phase is therefore a sort of topological homeostasis, reducing sharp gradients in shape to smoother topologies (Figure 6).
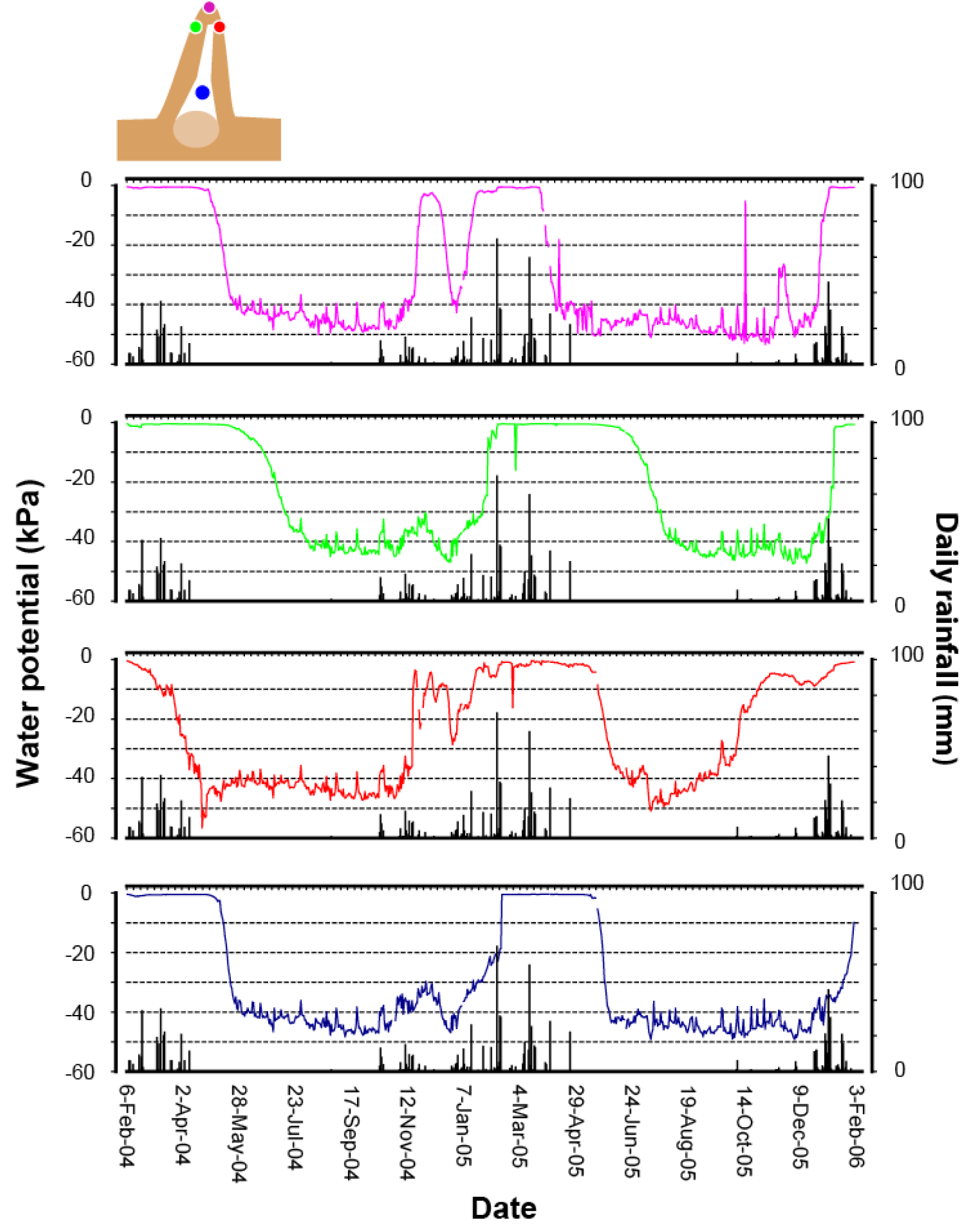
We are also discovering that these termites are avid regulators of nest moisture. Termites are generally intolerant of dry conditions, yet the Macrotermitinae are capable of inhabiting very dry habitats (Deshmukh, 1989). In Namibia, which is an arid country, Macrotermes can be found where annual rainfalls average as little as 250 mm. This is possible because these termites construct and regulate a moist nest environment (Turner, 2006). This alleviates the selective pressure on termites to adapt genetically to arid conditions, but there must be adaptation nevertheless. Placing a moist nest within a very dry environment drives substantial physical losses of water from the nest, which must be made good (Turner et al., 2007), Figure 8). During dry periods, the termites offset the physical losses of water by actively carrying water into the nest from deeper and moister soils. This is not a casual process: termites will go to great depths to obtain this water, as deep as a hundred meters or so by some anecdotal accounts (West, 1970). They also actively reconstruct the soil environment for several tens of meters around the nest, essentially building subsurface catchments that can gather water from sparse rainfalls and make it readily accessible to the termites (Boyer, 1969, 1971, 1973). The process is reversed during the rainy season. Following rainfall episodes, water percolates through the soil into the nest. Termites will work to offset this as well, carrying water in moist soil up out of the nest, into the mound, and ultimately to the mound surface where it can evaporate away (Turner et al., 2007). The end result is an impressive regulation of nest moisture throughout the year (Figure 9).
This homeostasis of nest moisture is coupled in an interesting way to the topological homeostasis that underscores mound remodeling. During the winter dry season, the termites apparently prefer to confine moisture
Figure?8
Water balance of a Macrotermes superorganism. Solid arrows signify work-driven water transport by termites. Dashed arrows represent passive movements of water due to infiltration from wet soils and evaporation through the mound.
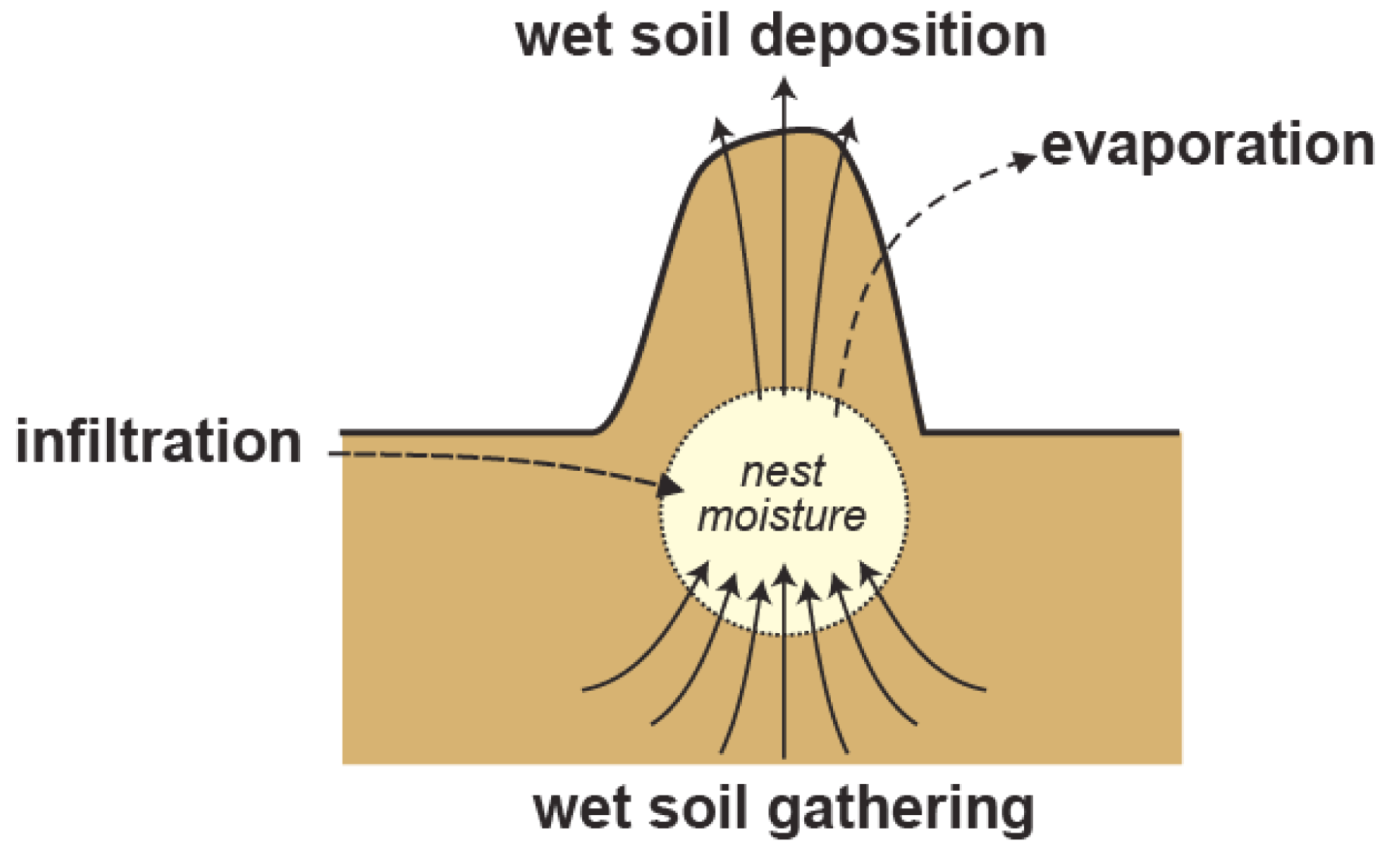{kind=link}
Figure?9
Moisture homeostasis in the nest of Macrotermes michaelseni. The moisture in the nest (blue trace) is maintained throughout the year, even as moisture in the adjacent soils (brown trace) dries considerably through the year. The center of the mound (green trace) is allowed to dry during the dry season, but becomes regulated during the rainy season. After Turner (2007).
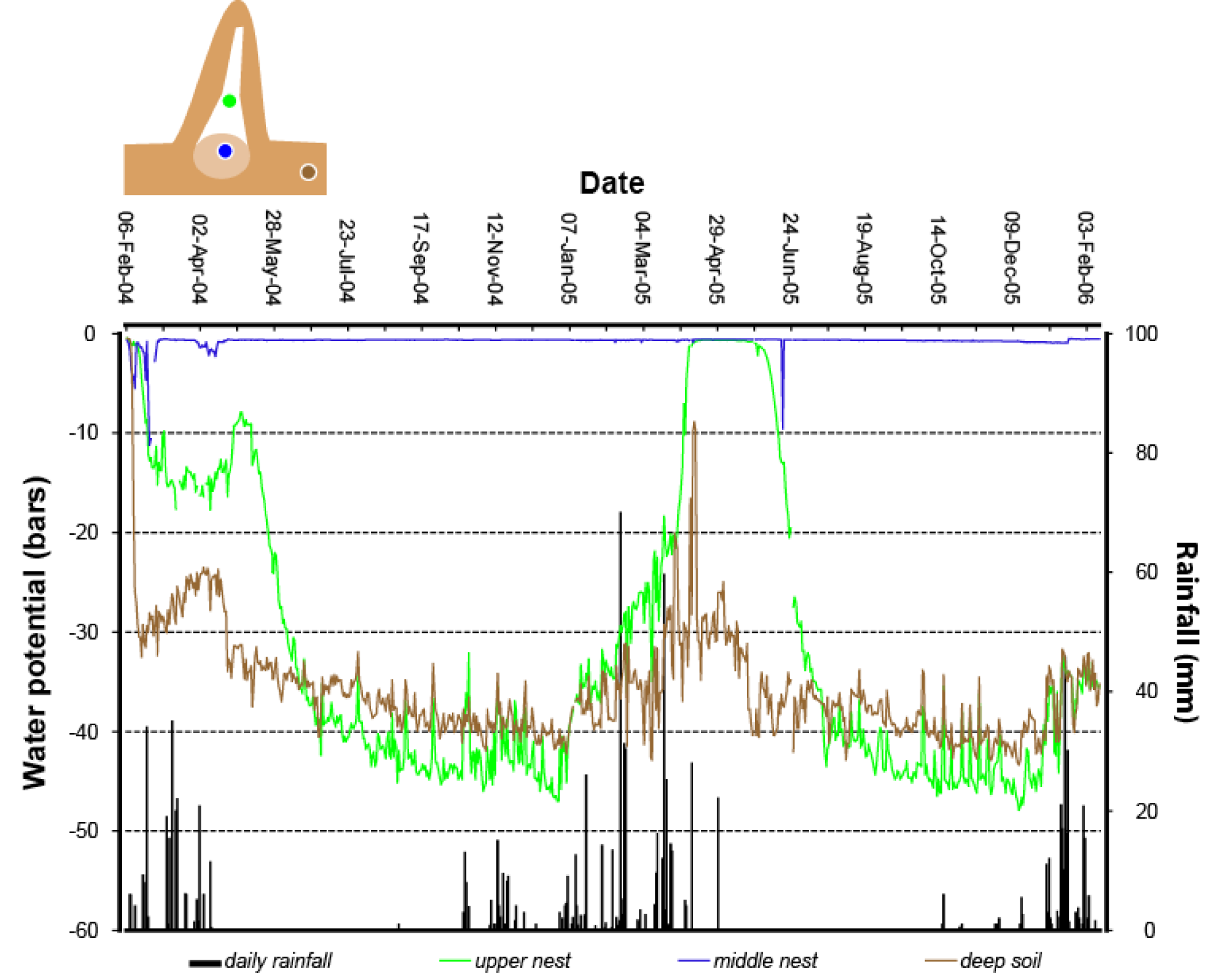{kind=link}
Is self-organization enough?
I could go on, but I hope my principal point is made: this system is the most impressive example of a superorganism of which I am aware. It exhibits coordination, integrity and a close matching of structure to function. It is also a complex system, with self-organized behaviors like stigmergy playing a foundational role in the emergence of these traits (Bonabeau et al., 1997). It is now apt to revisit the question I posed in my opening comments: is this
Figure?10
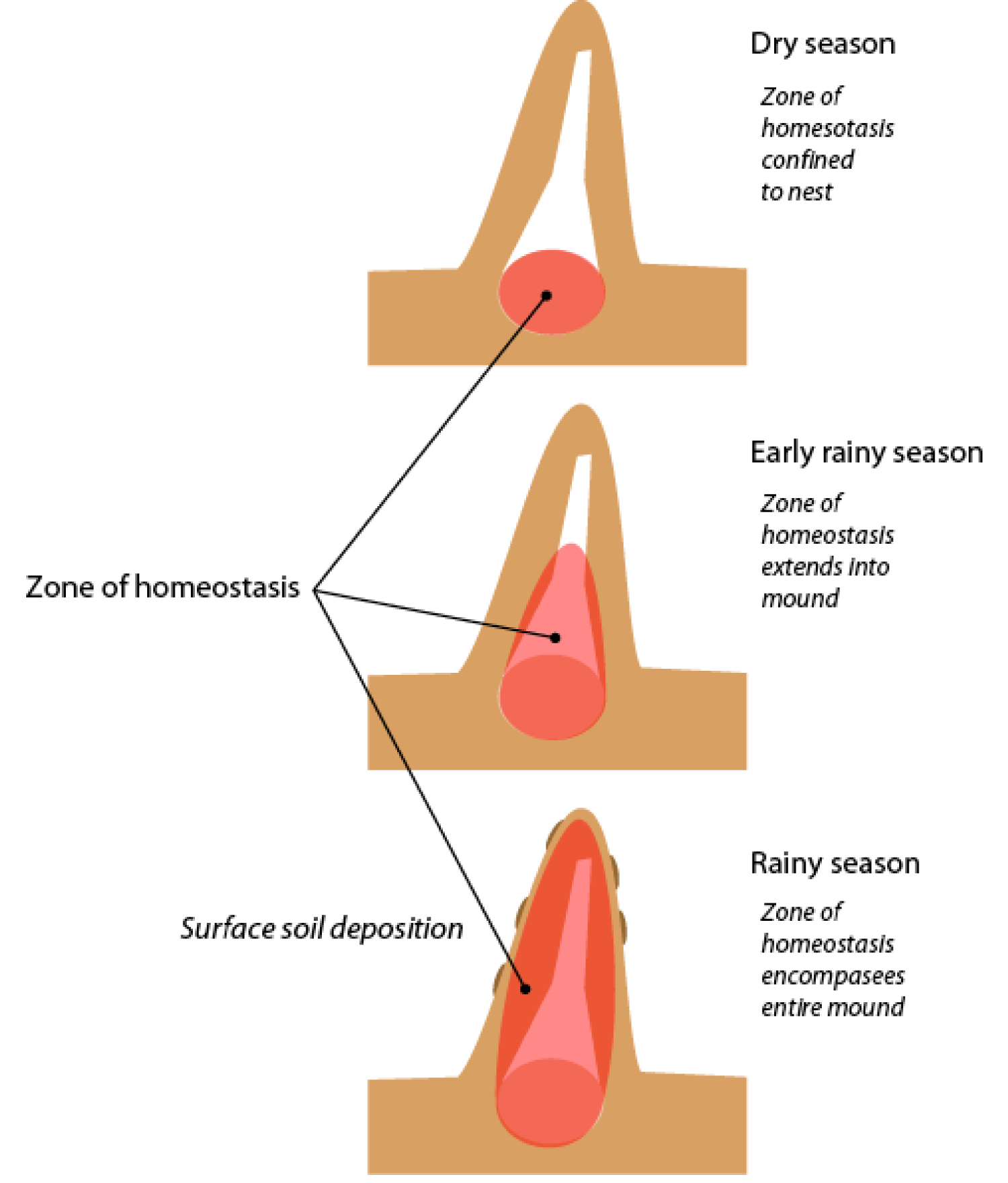
The expanding zone of moisture homeostasis in a Macrotermes michaelseni mound. As the rainy season proceeds, moisture throughout the mound comes to be regulated. After Turner (2007).
{kind=link}
My answer is “no”, and I justify my answer by a simple “natural experiment” the termites have obligingly provided. Self-organized behaviors like stigmergic building are not confined to the Macrotermitinae. They are, in fact, widespread among the termites. Yet, it is only among the Macrotermitinae, and from only a few genera among them, that the impressive superorganism I have described arises. The question therefore becomes: what, if not stigmergy,
Figure?11
The link between expanding zones of homeostasis and soil transport. Once the entire mound is enveloped in a zone of moisture homeostasis, soil deposition to the surface commences.
{kind=link}
As I have argued elsewhere (Turner, 2007), Macrotermes are but a very striking example of an ubiquitous, and unique, property of living systems: they comprise agents of homeostasis, whose modus operandi is to create new environments upon which homeostasis can be imposed. Among the termites, Macrotermes are certainly not unique in being agents of homeostasis. The unique Macrotermes superorganism emerges, however, when these agents are coupled to strong physiological drivers of matter and energy flux. I refer, of course, to the symbiotic fungi, whose collective metabolism exceeds the termites’ by a considerable factor. The mound is therefore properly regarded as the termites’ striving to keep ahead of the major environmental perturbation imposed by the nest’s fungal symbionts. This is aptly demonstrated by two phenomena, one that has played out over the evolution of fungiculture among the Macrotermitinae, and another that plays out over the life history of individual Macrotermes colonies.
The fungus-growing habit among the Macrotermitinae probably got its start as a hygienic measure. Fungi are usually serious parasites on the trove of cellulose that termites bring back to their colonies. Consequently, it is common for termites to store food in caches remote from the central nest. If a cache becomes infected with fungal parasites, it can be abandoned and isolated both from the colony and from other caches. The symbiosis between Macrotermes and Termitomyces probably began when the ancestors of Termitomyces proffered digestive benefits to the termites that parasitic fungi did not. The evolutionary trend among the Macrotermitinae has been to gather these “beneficial” caches together, presumably to protect them from infection by more aggressive parasitic fungal competitors. This trend culminates in the consolidated fungus gardens in the nest observed in the genera Macrotermes and Odontotermes.
With consolidation of the fungi, however, comes concentration of metabolic demand, producing nests with a high metabolic power density (energy release per unit nest volume; Turner, 2007). This profoundly affects the magnitude of fungus-driven perturbations of the nest environment. When fungal caches and the metabolic power they embody are widely dispersed, they experience little environmental perturbation: respiratory gases percolate readily through porous soil, and metabolic heat dissipates easily into the capacious thermal sink of the soil. With little environmental perturbation goes little impetus for the termites to remodel the environment of the cache. When the fungal caches are concentrated, however, heat and gases cannot so readily dissipate, resulting in more substantial perturbations of the nest environment. When these fungus-driven perturbations are coupled to the termites’ tendencies to remodel “uncomfortable” environments, the well-designed mound is the result.
This process can also be seen in the life history of individual Macrotermes colonies (Park and Turner, unpublished). Even in well-consolidated nests, termites often establish so-called accessory fungus gardens peripheral to the main nest. Why these accessory fungus gardens arise is unknown. However they arise, though, an accessory fungus garden represents a new, and peripheral, focus of high metabolic power density separate from the main nest. Remarkably, an accessory fungus garden invariably is associated with a “moundlet”, a new focus of intense upward transport of soil, driven by the same strong fungal perturbations that drive the construction of the principal mound. And they have the same consequence: construction of a designed “organ of physiology” to meet the metabolic demands of this new focus of metabolic power.
Whence the superorganism?
We now return to the question posed at the outset of this essay. One of life’s most striking attributes is the tendency of living agents to assemble into what we might call “organism-like” entities: cells into tissues, tissues into organs, organs into organisms, or organisms into superorganisms (Turner 2000, 2006b). Why should this be? Atomist doctrines, like self-organization or Neodarwinism, assert this tendency emerges spontaneously from simple agent-level interactions. As the Macrotermes superorganism clearly shows, these agent-level interactions are certainly important. By themselves, however, they fail to replicate the most interesting thing about the “organism-like” assemblage: its integrity, its cohesiveness, its coordination and its design. The missing element is a tendency—some might call it a “skyhook”, as Daniel Dennett provocatively puts it—that serves as an overarching organizing principle that is not derivable from the agents themselves. The missing element is homeostasis, which imparts to evolution and adaptation an inherent purposefulness that, I would argue, sits uncomfortably in the prevailing reductionism that pervades modern biology and to some extent, the emerging science of complexity.